
More Information
Submitted: August 16, 2023 | Approved: August 28, 2023 | Published: August 29, 2023
How to cite this article: Zhu YQ, Meng XY, Yang JH. Causal Link between Human Blood Metabolites and Asthma: An Investigation Using Mendelian Randomization. Arch Asthma Allergy Immunol. 2023; 7: 012-022.
DOI: 10.29328/journal.aaai.1001032
Copyright License: © 2023 Zhu YQ, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Keywords: Asthma; Serum metabolites; Mendelian randomization; Causal relationship; Risk Factors; 4-acetamidobutanoate
Abbreviations: MR: Mendelian Randomization; IVs: Instrumental Variables; IVW: Inverse Variance Weighted; SNPs: Single Nucleotide Polymorphisms; LD: Linkage Disequilibrium; β: Effect Size; SE: Standard Error; EAF: Effect Allele Frequency; R2: Instrumental Variable; N: Sample Size; k: SNP Count

Causal Link between Human Blood Metabolites and Asthma: An Investigation Using Mendelian Randomization
Yong-Qing Zhu1,2#, Xiao-Yan Meng1,2# and Jing-Hua Yang1,2*
1Clinical Systems Biology Laboratories, The First Affiliated Hospital of Zhengzhou University, Zhengzhou 450002, China
2The Academy of Medical Sciences, Zhengzhou University, Zhengzhou 450001, China
#These authors have contributed equally to this work and share first authorship
*Address for Correspondence: Jing-Hua Yang, Clinical Systems Biology Laboratories, The First Affiliated Hospital of Zhengzhou University, Zhengzhou 450002, China, Email: [email protected]
Background: Asthma, a chronic inflammatory respiratory ailment, is characterized by variable airflow obstruction and heightened bronchial reactivity. Despite therapeutic advancements, a comprehensive comprehension of its underlying metabolic mechanisms remains elusive. Metabolomics has emerged as a powerful approach to investigating the complex connections between serum metabolites and disease pathogenesis. However, exploring the causal relationship between serum metabolites and asthma susceptibility demands meticulous examination to unveil potential therapeutic targets.
Methods: Mendelian randomization (MR) approach was explored to investigate the potential causal associations between serum metabolites and asthma risk. The main analysis employed the inverse variance weighted method, supported by supplementary approaches such as MR-Egger, weighted median, weighted mode, and sample mode. To enhance the strength and credibility of our results, we conducted sensitivity analyses encompassing heterogeneity testing, assessment of horizontal pleiotropy, and leave-one-out analysis. Additionally, pathway enrichment analysis was performed to further elucidate the results.
Results: We identified 18 known and 12 unknown metabolites with potential associations with asthma risk. Among known metabolites, seven exhibited protective effects (e.g., 4-acetamidobutanoate, allantoin, kynurenine, oxidized bilirubin*), while eleven were considered risk factors (e.g., ornithine, N-acetylornithine, alanine). Through the integration of four additional MR models and sensitivity analyses, we revealed a connection between 4-acetamidobutanoate and approximately 6% lower asthma risk (OR = 0.94, 95% CI: 0.90–0.98).
Conclusions: Our MR analysis uncovered protective and risk-associated metabolites, alongside 12 unknown metabolites linked to asthma. Notably, 4-acetamidobutanoate demonstrated a nominal 6% reduction in asthma risk, highlighting its potential significance.
Asthma, a chronic inflammatory disorder of the airways, presents a substantial global health concern due to its prevalence, morbidity, and impact on individuals’ quality of life Asthma, characterized by airway inflammation and hyperresponsiveness, poses significant healthcare challenges worldwide [1]. Characterized by recurrent episodes of breathlessness, wheezing, chest tightness, and coughing, asthma affects millions of people worldwide, transcending age, gender, and geographic boundaries [2,3]. Effective therapies have greatly improved asthma morbidity and mortality over the past 15 years, but the precise etiology of asthma remains multifaceted and not fully elucidated [4]. It arises from intricate interactions between genetic predisposition and environmental triggers, resulting in airway inflammation, bronchoconstriction, and hypersensitivity responses [5]. With its variable and complex nature, asthma poses significant challenges in terms of diagnosis, treatment, and management [5]. As a result, a comprehensive understanding of the underlying mechanisms driving asthma’s onset, progression, and exacerbations is crucial for developing targeted interventions, improving patient outcomes, and alleviating the burden of this pervasive respiratory condition. This study aims to contribute to the ongoing exploration of asthma’s pathogenesis by investigating potential connections between serum metabolites and the risk of asthma using advanced analytical methodologies.
Metabolomics, a method to quantify small-molecule metabolites in biological samples, offers fresh perspectives on the connection between metabolic dysregulation and asthma [6]. Identifying unique metabolic patterns linked to asthma holds the potential for understanding disease variety and identifying new treatment targets. Increasing evidence points to metabolite alterations playing a role in asthma onset and worsening [7], highlighting the need for a thorough exploration of serum metabolisms’ association with asthma risk.
Mendelian Randomization (MR) is a statistical method that leverages genetic variants as instrumental variables to establish causal relationships between an exposure and an outcome in observational studies [8]. This method mimics randomized controlled trials by utilizing genetic variations as proxies for exposures, overcoming confounding and reverse causation typical in traditional observational studies [9]. The advantage lies in the natural randomization of exposure, akin to a randomized controlled trial [10], but without ethical complexities [11]. Additionally, MR can distinguish causation from correlation by assessing whether genetic variants impacting exposure also affect outcomes, shedding light on potential causal relationships [12]. MR’s value is evident in investigating hard-to-manipulate exposures, like risk factors such as smoking or alcohol consumption [13], to evaluate their impact on health outcomes [14]. Applying MR to study serum metabolites and asthma can distinguish whether specific metabolic pathways cause the disease or result from it [11]. This approach offers the potential to uncover intervention targets and strategies for asthma relief.
We have integrated MR as a foundational element in our investigation. Our primary goal is to explore the potential causal link between serum metabolites and asthma susceptibility. By leveraging genetic variation for causal inference, we aim to gain deeper insights into the complex interaction of metabolites and asthma.
Study design and date recourses
We conducted a two-sample MR investigation to explore the potential linkage between serum metabolites and asthma. This study was guided by three core assumptions (Figure 1) [8]. The first involves strong and direct relationships between IVs and exposure. The second requires IVs to have no connections with confounding factors. Lastly, IVs should solely impact the outcome through exposure pathways.
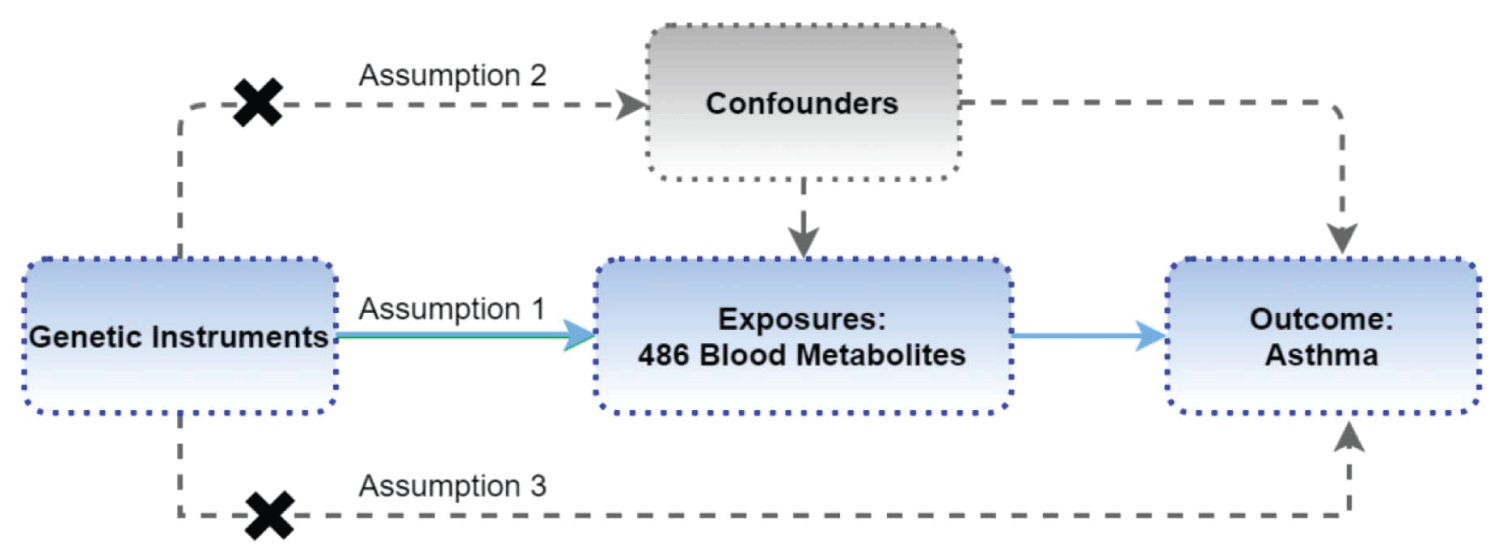
Figure 1: Study Design: Schematic overview of Two-Sample Mendelian Randomization Analyses in this study.
For the GWAS analysis of serum metabolites, data was sourced from the Metabolomics GWAS server (https://metabolomics.helmholtz-muenchen.de/gwas/), extracted from a comprehensive study by Shin, et al. [15]. The study cohort consisted of 7824 European adults who provided genetic samples [15]. Among over 2.1 million single nucleotide polymorphisms (SNPs), 486 metabolites underwent stringent quality assessments for analysis, encompassing 309 established metabolites and 177 unidentified ones. These 309 known metabolites were grouped into eight biochemical categories: amino acids, peptides, lipids, cofactors and vitamins, carbohydrates, energy-related compounds, nucleotides, and exotic substances.
As for asthma data, it was obtained from the IEU OpenGWAS project’s platform (https://gwas.mrcieu.ac.uk/), specifically dataset ukb-b-11297. The analysis encompassed 14283 asthma cases and 98300 controls of European descent, employing around 8.3 million SNPs for association evaluations.
Instrumental variable selection
For assumption (1), stringent screening identified IVs linked to blood metabolites. Due to a limited number of metabolite-associated SNPs, a slightly relaxed threshold (p < 1 × 10-5) was applied for SNP selection [16]. SNPs were grouped by eliminating Linkage Disequilibrium (LD) with R2 > 0.1 within 500 kb. To address weak instrument bias, each SNP underwent R2 and F statistic calculations based on parameters like effect size (β), Standard Error (SE), Effect Allele Frequency (EAF), instrumental variable (R2), sample size (N), and SNP count (k). SNPs with F statistic < 10 were excluded as inadequate instruments. Next, metabolite-associated SNPs were isolated from the outcome. Harmonization ensured consistency between exposure and outcome variables, addressing palindromic effects and allelic inconsistencies.
Formula 1:
Formula 2:
MR analysis
In this study, the primary two-sample MR analysis was conducted using the Inverse Variance Weighted (IVW) model [17], hinging upon pivotal assumptions—relevance, independence, exclusivity of IVs, and the genetic variation’s exclusive influence via exposure pathways. A confounding analysis of metabolites exhibiting IVW p < 0.05 revealed SNPs deviating from the MR Hypothesis. To ascertain IVs’ relationships with established risk factors (e.g., asthma, allergens, air pollution, etc.), the phenoscannerv2 website (http://www.phenoscanner.medschl.cam.ac.uk/) was utilized for IV metabolite investigation. Any SNPs showing associations with these confounders (p < 1 × 10-5) (Supplementary Table 1) were eliminated, along with outcome-related SNPs (p < 1 × 10-5) within IVs, ensuring autonomy and exclusivity. Metabolites displaying IVW p < 0.05 underwent subsequent MR Analysis for reinforced result dependability. Moreover, for an in-depth exploration of causal effects, we incorporated four additional MR Models: MR Egger regression, weighted median method, simple model-based estimator, and weighted model-based estimator [18].
| Table 1: Significant metabolites related to the risk of asthma according to IVW results (p < 0.05). | ||||||
| ID | Metabolite | nSNP | Beta | SE | p | OR (95%CI) |
| M01107 | allantoin | 19 | -0.03 | 0.01 | 2.11E-02 | 0.97(0.95-1.00) |
| M01493 | ornithine | 11 | 0.09 | 0.04 | 1.50E-02 | 1.09(1.02-1.17) |
| M01558 | 4-acetamidobutanoate | 40 | -0.06 | 0.02 | 5.81E-03 | 0.94(0.90-0.98) |
| M15140 | kynurenine | 39 | -0.07 | 0.03 | 9.79E-03 | 0.93(0.89-0.98) |
| M15630 | N-acetylornithine | 27 | 0.02 | 0.01 | 1.56E-02 | 1.02(1.00-1.03) |
| M15676 | 3-methyl-2-oxovalerate | 32 | 0.05 | 0.03 | 4.33E-02 | 1.06(1.00-1.11) |
| M18357 | glycylvaline | 7 | 0.03 | 0.01 | 2.00E-02 | 1.03(1.01-1.06) |
| M22116 | 4-methyl-2-oxopentanoate | 14 | 0.06 | 0.03 | 3.86E-02 | 1.07(1.00-1.13) |
| M32339 | alanine | 40 | 0.05 | 0.03 | 4.35E-02 | 1.05(1.00-1.11) |
| M32445 | 3-methylxanthine | 14 | 0.04 | 0.01 | 4.87E-03 | 1.04(1.01-1.06) |
| M32739 | X-11422--xanthine | 8 | 0.08 | 0.03 | 2.33E-02 | 1.08(1.01-1.16) |
| M33138 | X-11793--oxidized bilirubin* | 26 | -0.03 | 0.01 | 4.01E-03 | 0.97(0.95-0.99) |
| M33228 | 1-arachidonoylglycerophosphocholine* | 23 | 0.04 | 0.02 | 2.18E-02 | 1.04(1.01-1.08) |
| M34106 | bilirubin (E, Z or Z, E)* | 19 | -0.02 | 0.01 | 2.03E-02 | 0.98(0.96-1.00) |
| M34389 | 1-methylxanthine | 16 | 0.03 | 0.01 | 4.78E-02 | 1.03(1.00-1.05) |
| M34878 | X-13183--stearamide | 9 | -0.03 | 0.01 | 1.17E-02 | 0.97(0.94-0.99) |
| M35854 | threitol | 13 | 0.04 | 0.02 | 4.40E-02 | 1.04(1.00-1.08) |
| M36738 | gamma-glutamylglutamate | 10 | -0.03 | 0.01 | 4.34E-02 | 0.98(0.95-1.00) |
| M19415 | X-06351 | 4 | 0.08 | 0.04 | 4.15E-02 | 1.09(1.00-1.18) |
| M22032 | X-08766 | 16 | -0.05 | 0.02 | 2.33E-02 | 0.95(0.92-0.99) |
| M22548 | X-09026 | 15 | -0.06 | 0.03 | 4.05E-02 | 0.94(0.89-1.00) |
| M32518 | X-11204 | 42 | -0.07 | 0.03 | 1.54E-02 | 0.93(0.88-0.99) |
| M32761 | X-11444 | 19 | -0.04 | 0.02 | 4.09E-02 | 0.96(0.92-1.00) |
| M33195 | X-11850 | 8 | 0.02 | 0.01 | 3.30E-02 | 1.02(1.00-1.03) |
| M33751 | X-12329 | 20 | 0.01 | 0.00 | 3.28E-02 | 1.01(1.00-1.01) |
| M34516 | X-12833 | 13 | -0.01 | 0.00 | 3.94E-02 | 0.99(0.99-1.00) |
| M34530 | X-12847 | 8 | 0.02 | 0.01 | 3.17E-02 | 1.02(1.00-1.04) |
| M35978 | X-14057 | 18 | 0.03 | 0.02 | 2.45E-02 | 1.03(1.00-1.07) |
| M36585 | X-14658 | 11 | 0.03 | 0.01 | 4.18E-02 | 1.03(1.00-1.06) |
| M36673 | X-14745 | 13 | -0.05 | 0.02 | 4.04E-02 | 0.95(0.91-1.00) |
| Nsnp: Number of the SNP used for tests; SE: Standard error; OR: Odds ratio; 95% CI: 95% confidence interval; X: Unknown metabolite. | ||||||
Sensitivity analysis
Experimental variations, analytical platforms, and study subject diversity introduce potential heterogeneity in two-sample MR analyses, possibly causing biased causal effect estimates. To tackle this, we employed the Cochran Q test for heterogeneity assessment [19], where p < 0.05 signifies heterogeneity among IVs, while p > 0.05 indicates negligible impact on causal effect estimation.
The IVW method can be confounded by unknown factors and genetic multiplicity, introducing bias in causal effect estimates. To address this, we conducted a horizontal pleiotropy test by evaluating the MR-Egger regression intercept [20]. An intercept close to 0 (< 0.1) with p > 0.05 indicates no evidence of horizontal pleiotropy. We also used the MR-PRESSO method to assess horizontal pleiotropy and identify outliers.
After heterogeneity and horizontal pleiotropy tests, a sensitivity analysis was performed using the leave-one-out method [21] on qualified metabolites. This systematic approach removes each SNP, aggregates remaining SNPs to calculate the overall effect, and evaluates each SNP’s impact on metabolites. Stable overall error lines after SNP exclusion (all error lines not crossing 0) indicate reliable results.
Metabolic pathway and enrichment analysis
The analysis was conducted utilizing an online metabolomics data analysis platform (https://www.metaboanalyst.ca/MetaboAnalyst/faces/home.xhtml), specifically harnessing the Enrichment Analysis and Pathway Analysis modules within the Annotated Features mode. The corresponding IDs of these metabolites were subsequently extracted from the Human Metabolome Database (https://hmdb.ca/). Following this, the IDs were employed to interrogate pathways and enrichment using data derived from SMPDB (https://smpdb.ca/) and the KEGG database (https://www.kegg.jp/). This comprehensive strategy facilitated the aggregation of metabolite sets and pathways linked to the realm of asthma.
Statistical analysis
LD analyses utilized PLINK software (v1.9). Two-sample MR and sensitivity analyses employed the TwoSample MR package (v0.5.6) and GWASglue MR package (v0.0.0.9000) in R (v4.2.3).
MR analysis results
In this study, the IVW model served as the primary approach for estimating the causal relationships between blood metabolites and the risk of asthma (Supplementary Table 2).
| Table 2: Five MR models estimate the causal relationships between 18 known metabolites and the risk of asthma and tests for heterogeneity and horizontal pleiotropy. | ||||||
| Metabolite | Method | nSNP | p | OR 95%CI) | Pheter | Phoriz |
| allantoin | MR Egger | 19 | 4.07E-01 | 0.98(0.93-1.03) | 0.73 | 0.67 |
| Weighted median | 19 | 1.24E-01 | 0.97(0.94-1.01) | |||
| Inverse variance weighted | 19 | 2.11E-02 | 0.97(0.95-1.00) | 0.78 | ||
| Simple mode | 19 | 2.63E-01 | 0.97(0.91-1.02) | |||
| Weighted mode | 19 | 1.65E-01 | 0.97(0.94-1.01) | |||
| ornithine | MR Egger | 11 | 1.17E-01 | 1.21(0.97-1.51) | 0.46 | 0.34 |
| Weighted median | 11 | 6.45E-02 | 1.10(0.99-1.21) | |||
| Inverse variance weighted | 11 | 1.50E-02 | 1.09(1.02-1.17) | 0.46 | ||
| Simple mode | 11 | 3.53E-01 | 1.08(0.92-1.27) | |||
| Weighted mode | 11 | 1.82E-01 | 1.10(0.96-1.26) | |||
| 4-acetamidobutanoate | MR Egger | 40 | 7.34E-02 | 0.90(0.81-1.01) | 0.49 | 0.42 |
| Weighted median | 40 | 2.57E-03 | 0.90(0.85-0.97) | |||
| Inverse variance weighted | 40 | 5.81E-03 | 0.94(0.90-0.98) | 0.51 | ||
| Simple mode | 40 | 1.27E-01 | 0.90(0.78-1.03) | |||
| Weighted mode | 40 | 6.72E-03 | 0.89(0.82-0.96) | |||
| kynurenine | MR Egger | 39 | 1.98E-01 | 0.90(0.78-1.05) | 0.07 | 0.67 |
| Weighted median | 39 | 2.27E-01 | 0.96(0.89-1.03) | |||
| Inverse variance weighted | 39 | 9.79E-03 | 0.93(0.89-0.98) | 0.09 | ||
| Simple mode | 39 | 7.18E-01 | 0.97(0.84-1.13) | |||
| Weighted mode | 39 | 1.58E-01 | 0.92(0.82-1.03) | |||
| N-acetylornithine | MR Egger | 27 | 5.18E-02 | 1.02(1.00-1.05) | 0.28 | 0.54 |
| Weighted median | 27 | 4.81E-02 | 1.02(1.00-1.04) | |||
| Inverse variance weighted | 27 | 1.56E-02 | 1.02(1.00-1.03) | 0.30 | ||
| Simple mode | 27 | 4.39E-02 | 1.06(1.00-1.11) | |||
| Weighted mode | 27 | 5.50E-02 | 1.02(1.00-1.04) | |||
| 3-methyl-2-oxovalerate | MR Egger | 32 | 2.10E-01 | 1.13(0.94-1.36) | 0.50 | 0.46 |
| Weighted median | 32 | 2.23E-01 | 1.05(0.97-1.13) | |||
| Inverse variance weighted | 32 | 4.33E-02 | 1.06(1.00-1.11) | 0.52 | ||
| Simple mode | 32 | 3.96E-01 | 1.07(0.92-1.25) | |||
| Weighted mode | 32 | 2.25E-01 | 1.08(0.95-1.23) | |||
| glycylvaline | MR Egger | 7 | 7.41E-01 | 0.98(0.86-1.11) | 0.41 | 0.41 |
| Weighted median | 7 | 7.49E-02 | 1.04(1.00-1.08) | |||
| Inverse variance weighted | 7 | 2.00E-02 | 1.03(1.01-1.06) | 0.44 | ||
| Simple mode | 7 | 1.17E-01 | 1.06(1.00-1.13) | |||
| Weighted mode | 7 | 1.46E-01 | 1.05(0.99-1.12) | |||
| 4-methyl-2-oxopentanoate | MR Egger | 14 | 9.76E-01 | 1.00(1.82-1.23) | 0.63 | 0.56 |
| Weighted median | 14 | 1.31E-01 | 1.07(0.98-1.16) | |||
| Inverse variance weighted | 14 | 3.86E-02 | 1.07(1.00-1.13) | 0.68 | ||
| Simple mode | 14 | 3.20E-01 | 1.08(0.93-1.26) | |||
| Weighted mode | 14 | 1.77E-01 | 1.11(0.96-1.28) | |||
| alanine | MR Egger | 40 | 7.30E-01 | 0.97(0.79-1.18) | 0.64 | 0.38 |
| Weighted median | 40 | 1.21E-01 | 1.06(0.98-1.14) | |||
| Inverse variance weighted | 40 | 4.35E-02 | 1.05(1.00-1.11) | 0.65 | ||
| Simple mode | 40 | 6.05E-02 | 1.18(1.00-1.40) | |||
| Weighted mode | 40 | 8.58E-02 | 1.15(0.98-1.35) | |||
| 3-methylxanthine | MR Egger | 14 | 8.21E-01 | 0.99(0.93-1.06) | 0.46 | 0.19 |
| Weighted median | 14 | 1.35E-01 | 1.03(0.99-1.07) | |||
| Inverse variance weighted | 14 | 4.87E-03 | 1.04(1.01-1.06) | 0.39 | ||
| Simple mode | 14 | 2.88E-02 | 1.08(1.02-1.15) | |||
| Weighted mode | 14 | 4.99E-01 | 1.01(0.97-1.06) | |||
| X-11422--xanthine | MR Egger | 8 | 1.50E-01 | 1.16(0.97-1.39) | 0.96 | 0.42 |
| Weighted median | 8 | 2.14E-02 | 1.11(1.02-1.20) | |||
| Inverse variance weighted | 8 | 2.33E-02 | 1.08(1.01-1.16) | 0.94 | ||
| Simple mode | 8 | 1.38E-01 | 1.11(0.98-1.25) | |||
| Weighted mode | 8 | 1.10E-01 | 1.11(0.99-1.25) | |||
| X-11793--oxidized bilirubin* | MR Egger | 26 | 5.33E-02 | 0.96(0.92-1.00) | 0.31 | 0.51 |
| Weighted median | 26 | 4.69E-02 | 0.97(0.94-1.00) | |||
| Inverse variance weighted | 26 | 4.01E-03 | 0.97(0.95-0.99) | 0.34 | ||
| Simple mode | 26 | 2.71E-01 | 0.97(0.91-1.03) | |||
| Weighted mode | 26 | 5.53E-02 | 0.97(0.94-1.00) | |||
| 1-arachidonoylglycerophosphocholine* | MR Egger | 23 | 1.49E-01 | 1.06(0.98-1.13) | 0.27 | 0.65 |
| Weighted median | 23 | 1.19E-03 | 1.08(1.03-1.14) | |||
| Inverse variance weighted | 23 | 2.18E-02 | 1.04(1.01-1.08) | 0.31 | ||
| Simple mode | 23 | 9.02E-01 | 0.99(0.89-1.10) | |||
| Weighted mode | 23 | 1.32E-02 | 1.08(1.02-1.13) | |||
| bilirubin (E, Z or Z, E)* | MR Egger | 19 | 1.44E-02 | 0.94(0.90-0.98) | 0.67 | 0.07 |
| Weighted median | 19 | 2.52E-02 | 0.97(0.94-1.00) | |||
| Inverse variance weighted | 19 | 2.03E-02 | 0.98(0.96-1.00) | 0.48 | ||
| Simple mode | 19 | 7.72E-02 | 0.95(0.91-1.00) | |||
| Weighted mode | 19 | 3.52E-02 | 0.96(0.93-0.99) | |||
| 1-methylxanthine | MR Egger | 16 | 4.14E-02 | 1.05(1.01-1.11) | 1.00 | 0.18 |
| Weighted median | 16 | 5.15E-02 | 1.03(1.00-1.07) | |||
| Inverse variance weighted | 16 | 4.78E-02 | 1.03(1.00-1.05) | 0.98 | ||
| Simple mode | 16 | 3.66E-01 | 1.03(0.97-1.09) | |||
| Weighted mode | 16 | 8.42E-02 | 1.03(1.00-1.07) | |||
| X-13183--stearamide | MR Egger | 9 | 3.51E-02 | 0.92(0.87-0.98) | 0.72 | 0.14 |
| Weighted median | 9 | 3.89E-01 | 0.98(0.94-1.02) | |||
| Inverse variance weighted | 9 | 1.17E-02 | 0.97(0.94-0.99) | 0.50 | ||
| Simple mode | 9 | 6.08E-01 | 0.98(0.92-1.05) | |||
| Weighted mode | 9 | 5.87E-01 | 0.98(0.92-1.04) | |||
| threitol | MR Egger | 13 | 8.89E-01 | 1.01(0.92-1.10) | 0.37 | 0.46 |
| Weighted median | 13 | 2.55E-01 | 1.03(0.98-1.08) | |||
| Inverse variance weighted | 13 | 4.40E-02 | 1.04(1.00-1.08) | 0.40 | ||
| Simple mode | 13 | 4.76E-01 | 1.03(0.96-1.11) | |||
| Weighted mode | 13 | 4.00E-01 | 1.03(0.97-1.09) | |||
| gamma-glutamylglutamate | MR Egger | 10 | 4.39E-02 | 0.93(0.88-0.99) | 0.49 | 0.13 |
| Weighted median | 10 | 1.20E-01 | 0.98(0.95-1.01) | |||
| Inverse variance weighted | 10 | 4.34E-02 | 0.98(0.95-1.00) | 0.32 | ||
| Simple mode | 10 | 5.57E-01 | 0.99(0.94-1.03) | |||
| Weighted mode | 10 | 9.26E-02 | 0.97(0.93-1.00) | |||
| Notes: nSNP, number of the SNP used for tests; OR, odds ratio; 95% CI, 95% confidence interval; Pheter: PHeterogeneity; Phoriz: PHorizontal pleiotropy. | ||||||
A total of 30 metabolites comprising 18 known metabolites and 12 unknown metabolites displayed a significant relationship (p < 0.05, IVW method) with asthma risk (Table 1). The 18 known metabolites could be classified into distinct categories: Amino acids (ornithine, 4-acetamidobutanoate, Kynurenine, N-acetylornithine, 3-methyl-2-oxovalerate, 4-methyl-2-oxopentanoate, alanine), Cofactors and vitamins (X-11793--oxidized bilirubin*, bilirubin (E, Z or Z, E)*), Lipids (1-arachidonoylglycerophosphocholine*, X-13183--stearamide), Nucleotides (allantoin, X-11422--xanthine), Peptides (glycylvaline, gamma-glutamylglutamate), Xenobiotics (3-methylxanthine, 1-methylxanthine), and Carbohydrates (threitol).
These known metabolites can be classified into 11 risk factors and 7 protective metabolites, based on their connection with asthma risk through the IVW method (Table 2). Of these, allantoin (OR = 0.97, 95% CI: 0.95-1.00), 4-acetamidobutanoate (OR = 0.94, 95% CI: 0.90-0.98), kynurenine (OR = 0.93, 95% CI: 0.89-0.98), X-11793--oxidized bilirubin* (OR = 0.97, 95% CI: 0.95-0.99), bilirubin (E, Z or Z, E)* (OR = 0.98, 95% CI: 0.96-1.00), X-13183--stearamide (OR = 0.97, 95% CI: 0.94-0.99) and gamma-glutamylglutamate (OR = 0.98, 95% CI: 0.95-1.00) exhibited potential reductions in asthma risk (Table 2). Conversely, ornithine (OR = 1.09, 95% CI: 1.02-1.17), N-acetylornithine (OR = 1.02, 95% CI:1.00-1.03), 3-methyl-2-oxovalerate (OR = 1.06, 95% CI: 1.00-1.11), glycylvaline (OR = 1.03, 95% CI: 1.01-1.06), 4-methyl-2-oxopentanoate (OR = 1.07, 95%CI: 1.00-1.13), alanine (OR =1.05, 95% CI: 1.00-1.11), 3-methylxanthine (OR = 1.04, 95%CI: 1.01-1.06), X-11422--xanthine (OR = 1.08, 95% CI: 1.01-1.16),1-arachidonoylglycerophosphocholine* (OR = 1.04, 95% CI: 1.01-1.08), 1-methylxanthine(OR = 1.03, 95% CI: 1.00-1.05) and threitol(OR = 1.04, 95% CI: 1.00-1.08) displayed potential increased asthma risk (Table 2).
Subsequently, we deployed an ensemble of four supplementary models (Bowden, et al. 2015) for the meticulous assessment of causal effects between these metabolites and the risk of asthma (Table 2). Three consistently demonstrated significance in a minimum of three MR models and consistently underscored the causal implications across the entire spectrum of models (Table 2). The three metabolites are 4-acetamidobutanoate (P IVW = 5.81 × 10−3, P MR Egger = 7.34 × 10−2, P Weighted median = 2.57 × 10−3, P Simple mode = 1.27 × 10−1, P Weighted mode = 6.72 × 10−3), N-acetylornithine (P IVW = 1.56 × 10−2, P MR Egger = 5.18 × 10−2, P Weighted median = 4.81 × 10−2, P Simple mode = 4.39 × 10−2, P Weighted mode = 5.50 × 10−2), and bilirubin (E, Z or Z, E)* (P IVW = 2.03 × 10−2, P MR Egger = 1.44 × 10−2, P Weighted median = 2.52 × 10−2, P Simple mode = 7.72 × 10−2, P Weighted mode = 3.52 × 10−2) (Table 2). The congruence is manifested in the outcomes for 4-acetamidobutanoate (Figure 2A), N-acetylornithine (Figure 3A), and bilirubin (E, Z or Z, E) (Figure 4A) across all five models.
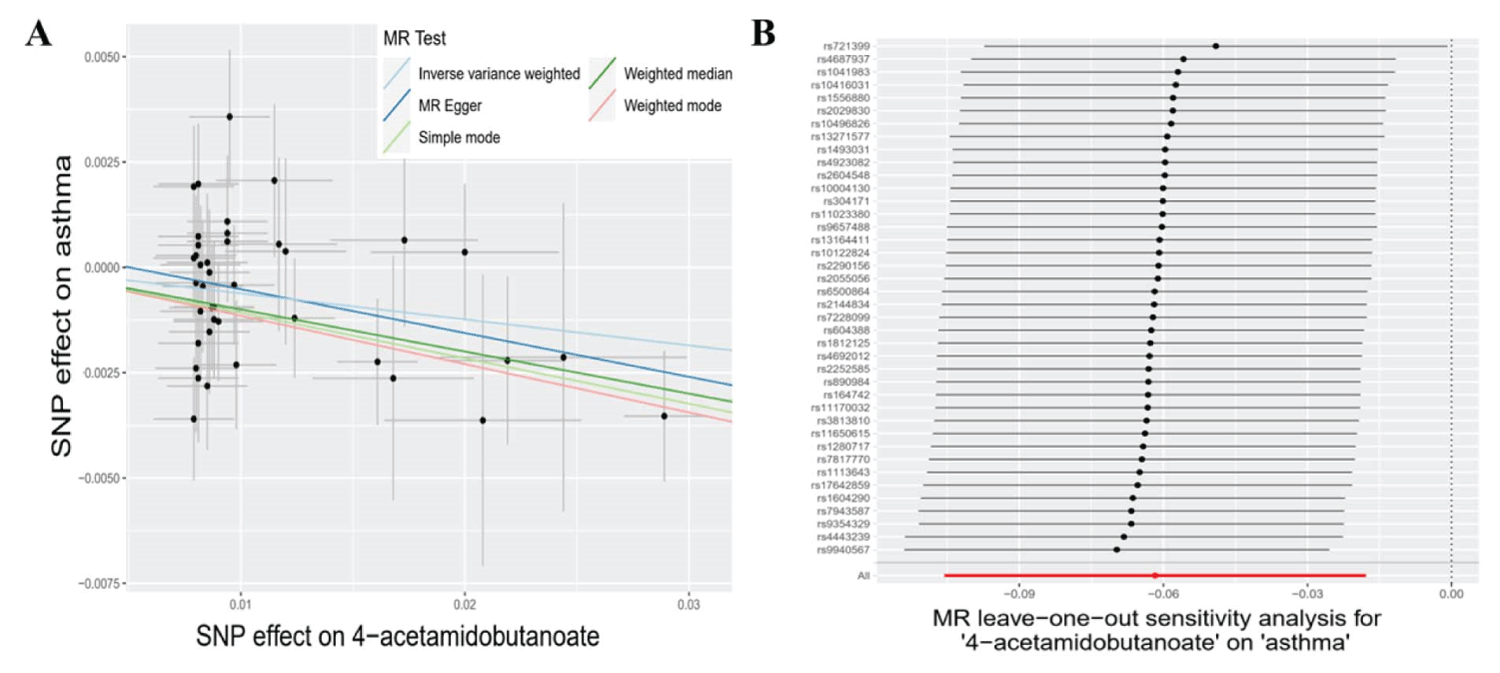
Figure 2: The causal relationships between 4-acetamidobutanoate and asthma, along with sensitivity analyses. A: Scatter plots of the 5 MR models for 4-acetamidobutanoate with the risk of asthma. light blue line: inverse variance weighted; blue line: MR Egger; light green line: simple model-based estimator; green line: weighted median estimator; red line: weighted model-based estimator. B: Forest plots show the results of leave-one-out analyses of 4-acetamidobutanoate.
Assessment of the reliability and stability of the results
To ensure the credibility of our findings, we subjected the reliability and consistency of outcomes pertaining to the known metabolites to stringent examinations. Employing MR-Egger and MR-PRESSO methods, the results demonstrated p - values > 0.05. Furthermore, the nearly null intercept of the MR-Egger regression (< 0.1) signaled the absence of heterogeneity and horizontal pleiotropy within these metabolites (Table 2 and Supplementary Table 3).
| Table 3: Re-analyses results of five MR models after the removal of sensitive SNP for N-acetylornithine and bilirubin (E, Z or Z, E). | ||||
| Metabolite | Method | nSNP | p | OR (95%CI) |
| MR Egger | 26 | 1.08E-01 | 1.02(1.00-1.05) | |
| Weighted median | 26 | 1.26E-01 | 1.02(1.00-1.03) | |
| N-acetylornithine | Inverse variance weighted | 26 | 5.28E-02 | 1.02(1.00-1.03) |
| Simple mode | 26 | 2.07E-01 | 0.96(0.89-1.02) | |
| Weighted mode | 26 | 1.13E-01 | 1.02(1.00-1.04) | |
| MR Egger | 17 | 9.13E-02 | 0.94(0.88-1.01) | |
| Weighted median | 17 | 1.94E-01 | 0.98(0.94-1.01) | |
| bilirubin (E,Z or Z,E)* | Inverse variance weighted | 17 | 2.39E-01 | 0.99(0.96-1.01) |
| Simple mode | 17 | 1.85E-01 | 0.95(0.89-1.02) | |
| Weighted mode | 17 | 1.48E-01 | 0.95(0.89-1.01) | |
| Notes: nSNP, number of the SNP used for tests; OR, odds ratio; 95% CI, 95% confidence interval. | ||||
Concerning the three metabolites, 4-acetamidobutanoate, N-acetylornithine, and bilirubin (E, Z or Z, E)*, which showcased remarkable resilience by consistently revealing significance in a minimum of three MR models, we executed sensitivity analyses utilizing a leave-one-out approach to scrutinize their stability. The outcomes revealed that all SNPs linked with 4-acetamidobutanoate exhibited insensitivity to the results (Figure 2B), affirming a steadfast and noteworthy 6% reduction in asthma risk (Table 2, Figure 5). Conversely, it came to light that a solitary instrumental variable (rs7594485) associated with N-acetylornithine (Figure 3B), and two IVs (rs887829 and rs28900385) linked with bilirubin (E, Z or Z, E)* (Figure 4B), exerted considerable influence on the outcome. Subsequently, upon exclusion of rs7594485, rs887829, and rs28900385, we proceeded to re-conduct MR analyses through the five models, yielding results of N-acetylornithine and bilirubin (E, Z or Z, E)* that no longer retained significance (Table 3).
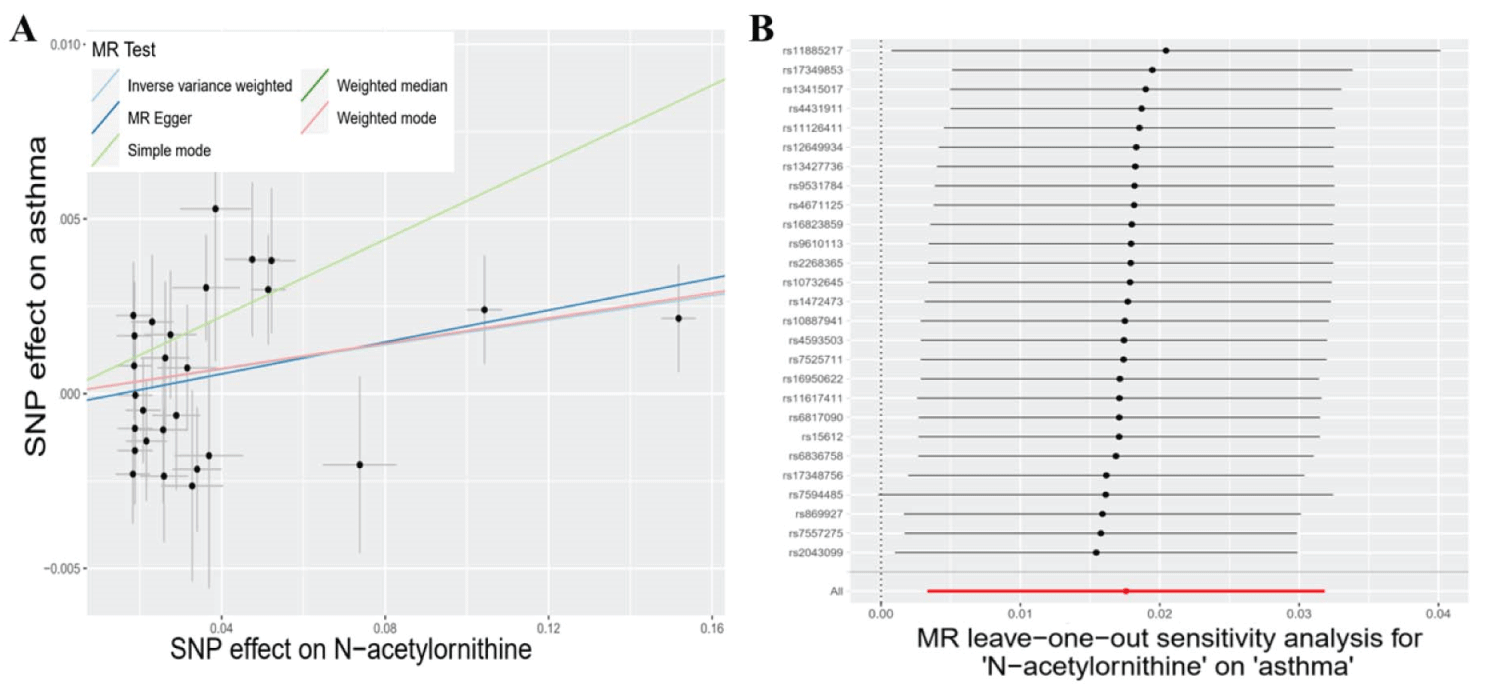
Figure 3: The causal associations between N-acetylornithine and asthma, followed by in-depth sensitivity analyses. A: Illustrations of scatter plots for N-acetylornithine's association with asthma risk across the 5 MR models. The light blue line represents inverse variance weighted, blue for MR Egger, light green for the simple model-based estimator, green for the weighted median estimator, and red for the weighted model-based estimator. B: Forest plots visually display the outcomes from leave-one-out analyses for N-acetylornithine.
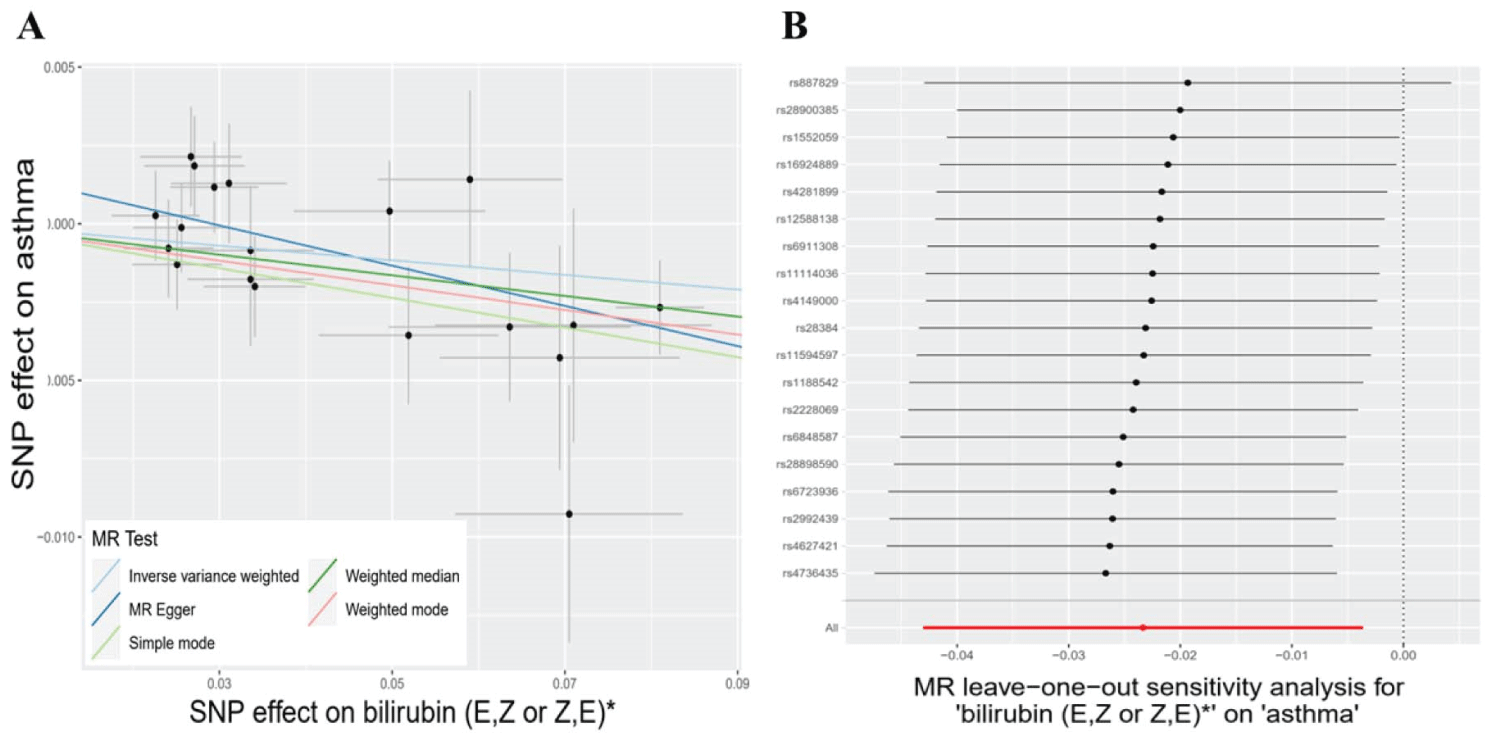
Figure 4: The causal relationships between bilirubin (E, Z or Z, E) and asthma, and their subsequent sensitivity analyses. A: The risk of asthma is depicted in scatter plots across 5 MR models for bilirubin (E, Z or Z, E). The associations are represented by different lines: light blue for inverse variance weighted, blue for MR Egger, light green for the simple model-based estimator, green for the weighted median estimator, and red for the weighted model-based estimator. B: Leave-one-out analyses of bilirubin (E, Z or Z, E) are displayed in forest plots, providing insights into the results.
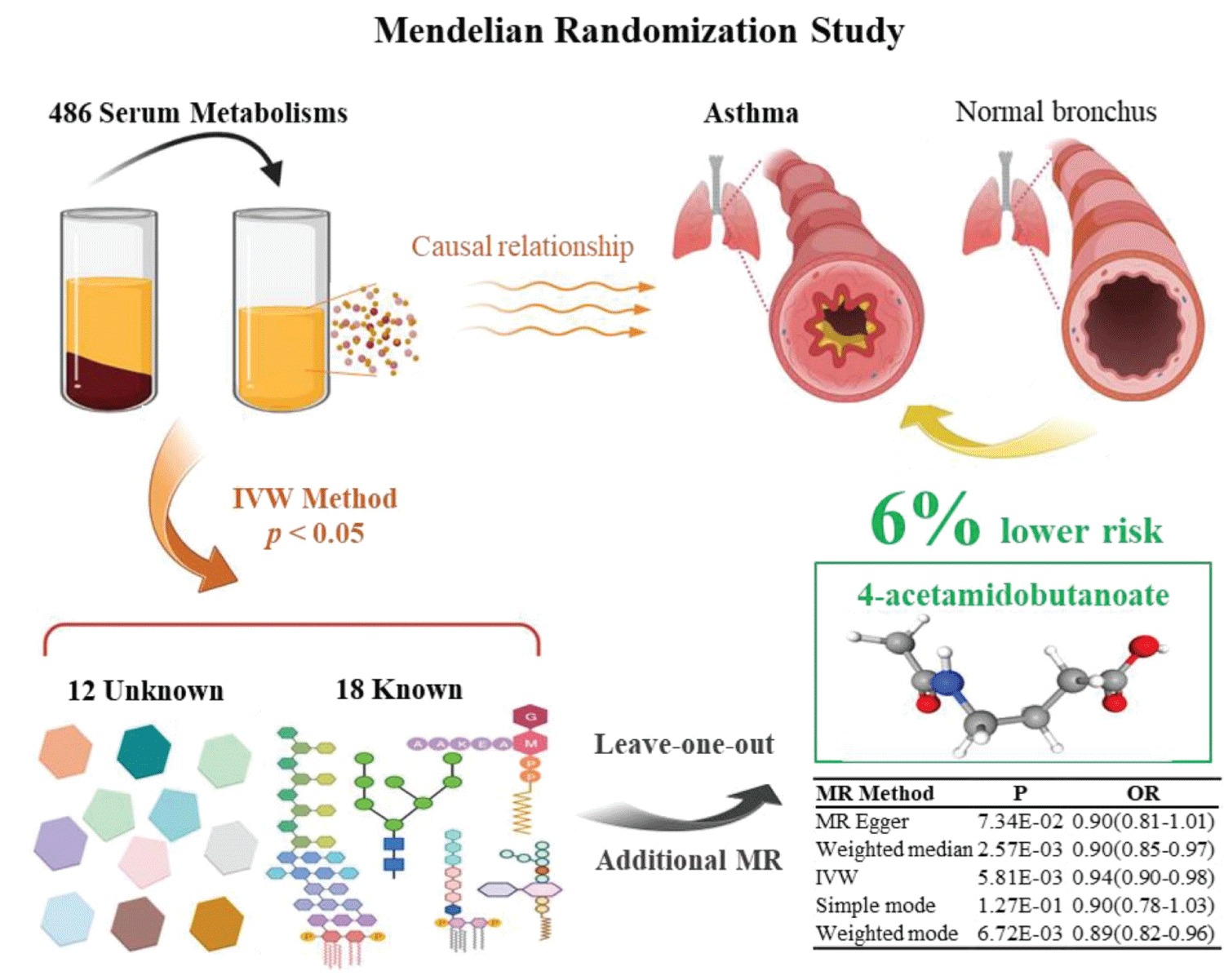
Figure 5: Graphical summary of the Mendelian Randomization Study.
Metabolic pathway and enrichment analysis
In the pathway analysis of known metabolites, we uncovered five metabolic pathways that exhibited relative significance (p < 0.05). The identified metabolic pathways encompassed “ Valine, leucine and isoleucine biosynthesis” (p = 0.001), “Arginine biosynthesis” (p = 0.004), Arginine and proline metabolism (p = 0.028), and “ Valine, leucine and isoleucine degradation” (p = 0.031) (Supplementary Figure 1, and Supplementary Tables 4,5). As for the enrichment analysis, the identified metabolic pathways encompassed the “Urea Cycle” (p = 0.025) and “Porphyrin Metabolism” (p = 0.045) (Supplementary Figures 2-3, and Supplementary Table 6).
Asthma pathogenesis involves complex interactions between genetic, environmental [22], and immunological factors [23,24]. The noteworthy interest in metabolic dysregulation’s role in asthma susceptibility and progression has grown [25-27]. Metabolomic studies have unveiled identifiable metabolic patterns associated with asthma. Altered metabolite levels in asthma individuals, compared to healthy controls, suggest a credible link between metabolic disruptions and disease expression [28]. The clinical implications of metabolism’s importance in asthma are profound. Metabolomic profiling offers avenues for identifying crucial biomarkers, essential for asthma diagnosis, severity assessment, and treatment response prediction.
In this study, we carried out an impartial two-sample MR analysis aimed at investigating the potential causal link between 486 blood metabolites and the susceptibility to asthma. To fortify the rigor of our inquiry, we meticulously collected the most extensive Genome-Wide Association Study (GWAS) data and comprehensive asthma GWAS summary data from publicly accessible databases. By leveraging genetic variants as IVs, we discerned 18 established and 12 uncharacterized metabolites that exhibited promise as potential predictors of asthma risk, as elucidated by our primary IVW analysis. Within this group of recognized metabolites, they can be compartmentalized into seven factors of protection (allantoin, 4-acetamidobutanoate, kynurenine, X-11793--oxidized bilirubin*, bilirubin (E, Z or Z, E), X-13183—stearamide, and gamma-glutamylglutamate) and eleven factors of risk (ornithine, N-acetylornithine, 3-methyl-2-oxovalerate, glycylvaline, 4-methyl-2-oxopentanoate, alanine, 3-methylxanthine, X-11422--xanthine, 1-arachidonoylglycerophosphocholine*, 1-methylxanthine, and threitol). Through the metabolic pathway analysis of the selected known metabolites, they were concentrated in the metabolic pathways related to urea metabolism and arginine metabolism, suggesting that metabolites may play a certain role in the pathogenesis and progression of asthma through these pathways.
To augment the reliability and stability of our findings, we employed supplementary MR models. The outcomes uniformly substantiated the correlation between 4-acetamidobutanoate, N-acetylornithine, and bilirubin (E, Z or Z, E)* with a reduced asthma risk across a minimum of three MR models. Nonetheless, prudence is advised when construing the causal link involving N-acetylornithine and bilirubin (E, Z or Z, E)*, given that they faltered in the concluding leave-one-out analysis, necessitating further investigation.
Also recognized as 4-acetamidobutanoic acid or N-acetyl-4-aminobutyric acid, 4-acetamidobutanoate belongs to the category of gamma amino acids and derivatives [29]. It emerges as a byproduct of the urea cycle and the metabolism of amino groups, while also originating from NAD-linked aldehyde [30]. In our study, through metabolic pathway enrichment analysis, we found that 4-acetamidobutanoate is involved in the Urea Cycle, and the metabolism of valine, leucine, isoleucine, and arginine. The urea cycle, critical for detoxifying ammonia, sparks interest in its potential asthma link [31]. Recent research highlights its role in immune response and inflammation modulation, central to asthma [32]. Ammonia, a byproduct, impacts airway muscle contraction and bronchoconstriction, pivotal in asthma features [33]. Valine, leucine, and isoleucine, termed branched-chain amino acids, have emerged in asthma discussions [34]. Studies hint at associations between altered amino acid levels and asthma susceptibility or severity [35]. Arginine, a semi-essential amino acid, gains attention for its possible asthma role [36]. It affects immune responses and airway function, vital in asthma development [37]. Arginine metabolism produces Nitric Oxide (NO), influencing airway smooth muscle tone and inflammation [38]. Elevated NO levels in asthma connect arginine to bronchial constriction and symptoms [39].
4-acetamidobutanoate presence extends across all eukaryotic organisms, spanning from yeast to humans, and is detectable in various food items like blackberry, cassava, pepper, and napa cabbage [40]. Although speculative, it is plausible that the consumption of these foods might have the potential to mitigate asthma incidence. Nonetheless, the precise causal correlation between 4-acetamidobutanoate and asthma remains partially apprehended. Despite this, our pioneering findings unveil an extraordinary revelation: 4-acetamidobutanoate exhibits a substantial connection, linked to a noteworthy 6% reduction in asthma incidence.
Besides serum metabolites, various factors influence asthma risk, including lifestyle choices such as smoking [41]and alcohol use [42], which heighten susceptibility and exacerbations. Obesity also increases asthma risk due to inflammation and respiratory effects [42]. Puberty timing, particularly in females, impacts asthma via hormonal shifts affecting airway responsiveness [43]. Environmental factors like allergens, pollution, and infections contribute. Genetic predisposition, observed in familial asthma cases, is significant [44]. Prenatal factors like maternal smoking and allergen exposure elevate offspring asthma chances [45]. These multifaceted influences involve lifestyle, environment, genetics, and hormones, molding asthma’s development and severity. Recognizing these complexities is also vital for comprehensive asthma prevention and management approaches.
Our study introduces significant innovations. Firstly, we adopt a molecular mechanism perspective, treating blood metabolites as exposure factors. This approach establishes a robust theoretical foundation and holds clinical research value in probing causal connections between metabolites and asthma risk. Secondly, our rigorous commitment to high-quality control, diverse methodologies, and multiple analytical approaches ensures comprehensive evaluation of causal effects, guaranteeing the reliability and stability of our findings. Thirdly, unlike prior MR analyses focused on individual exposures, our thorough examination of numerous blood metabolites presents substantial analytical challenges. Our proposed analytical strategy offers valuable insights for comparable investigations.
However, we acknowledge limitations. Two-fifths of the asthma risk predictors identified, using the IVW method, are unidentified metabolites with uncertain functional profiles, limiting the scope of our findings. While a nominal causal link between 4-acetamidobutanoate and asthma is evident through our unbiased two-sample MR approach, this relationship remains theoretical, pending mechanistic validation. Thus, further inquiry is essential to clarify 4-acetamidobutanoate’s role in asthma pathogenesis and establish a conclusive confirmation of this causal connection. By embracing these limitations as avenues for growth, our study provides a foundation for future research, enriching our understanding of asthma and its molecular basis.
We utilized a two-sample MR approach to uncover causal links between 486 blood metabolites and asthma in a vast cohort of over 0.11 million individuals of European descent. Through meticulous analysis, we identified 30 serum metabolites associated with asthma, comprising 7 protective metabolites, 11 risk factors, and 12 previously unknown metabolites. Notably, our findings suggest a 6% reduction in asthma risk attributed to 4-acetamidobutanoate. These revelations significantly enhance our grasp of the intricate interplay between blood metabolites and asthma, offering the potential for personalized insights or markers that elucidate biological variations in disease status. By illuminating these crucial connections, our study paves the way for future research avenues, propelling advancements in asthma prevention, diagnosis, and management, and ultimately fostering improved global lung health and well-being.
Author contributions
Yong-Qing Zhu oversaw data acquisition, performed MR analysis, and organized results. The article’s co-authors are Yong-Qing Zhu and Xiao-Yan Meng. Jing-Hua Yang guided students, contributed to idea development, and supervised the study. All authors engaged in active project discussions.
Funding
This work was partially supported by grants to Jing-Hua Yang from the National Natural Science Foundation of China (#32070157), the Department of Science and Technology of Henan Province (#201400210500), and a Tai-Shan Scholarship with Xinfa Pharmaceutical of Shandong Province.
Supplementary material: Supplementary material is available online in the Supporting Information section.
Supplementary material is available online in the Supporting Information section.
We extend our gratitude to Shin, et al. and Huyghe, et al. for sharing the original GWAS data, as well as to the GWAS Catalog database.
Ethics statement
Our sincere thanks go to Shin, et al. and Huyghe, et al. for generously providing the original GWAS data, along with our appreciation for the GWAS Catalog database.\
- Porsbjerg C, Melén E, Lehtimäki L, Shaw D. Asthma. Lancet. 2023 Mar 11;401(10379):858-873. doi: 10.1016/S0140-6736(22)02125-0. Epub 2023 Jan 19. PMID: 36682372.
- Chowdhury NU, Guntur VP, Newcomb DC, Wechsler ME. Sex and gender in asthma. Eur Respir Rev. 2021 Nov 17;30(162):210067. doi: 10.1183/16000617.0067-2021. PMID: 34789462; PMCID: PMC8783601.
- Stern J, Pier J, Litonjua AA. Asthma epidemiology and risk factors. Semin Immunopathol. 2020 Feb;42(1):5-15. doi: 10.1007/s00281-020-00785-1. Epub 2020 Feb 4. PMID: 32020334.
- Zhou L, Hao M, Fan X, Lao Z, Li M, Shang E. Effects of Houpo Mahuang Decoction on serum metabolism and TRPV1/Ca2+/TJs in asthma. J Ethnopharmacol. 2023 Feb 10;302(Pt A):115873. doi: 10.1016/j.jep.2022.115873. Epub 2022 Oct 26. PMID: 36309114.
- Agache I, Eguiluz-Gracia I, Cojanu C, Laculiceanu A, Del Giacco S, Zemelka-Wiacek M, Kosowska A, Akdis CA, Jutel M. Advances and highlights in asthma in 2021. Allergy. 2021 Nov;76(11):3390-3407. doi: 10.1111/all.15054. Epub 2021 Aug 29. PMID: 34392546.
- Liang Y, Gai XY, Chang C, Zhang X, Wang J, Li TT. Metabolomic Profiling Differences among Asthma, COPD, and Healthy Subjects: A LC-MS-based Metabolomic Analysis. Biomed Environ Sci. 2019 Sep;32(9):659-672. doi: 10.3967/bes2019.085. PMID: 31635682.
- Dixon AE, Holguin F. Diet and Metabolism in the Evolution of Asthma and Obesity. Clin Chest Med. 2019 Mar;40(1):97-106. doi: 10.1016/j.ccm.2018.10.007. Epub 2018 Dec 19. PMID: 30691720; PMCID: PMC6355154.
- Emdin CA, Khera AV, Kathiresan S. Mendelian Randomization. JAMA. 2017 Nov 21;318(19):1925-1926. doi: 10.1001/jama.2017.17219. PMID: 29164242.
- Bowden J, Holmes MV. Meta-analysis and Mendelian randomization: A review. Res Synth Methods. 2019 Dec;10(4):486-496. doi: 10.1002/jrsm.1346. Epub 2019 Apr 23. PMID: 30861319; PMCID: PMC6973275.
- Birney E. Mendelian Randomization. Cold Spring Harb Perspect Med. 2022 May 17;12(4):a041302. doi: 10.1101/cshperspect.a041302. PMID: 34872952; PMCID: PMC9121891.
- Sekula P, Del Greco M F, Pattaro C, Köttgen A. Mendelian Randomization as an Approach to Assess Causality Using Observational Data. J Am Soc Nephrol. 2016 Nov;27(11):3253-3265. doi: 10.1681/ASN.2016010098. Epub 2016 Aug 2. PMID: 27486138; PMCID: PMC5084898.
- Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014 Sep 15;23(R1):R89-98. doi: 10.1093/hmg/ddu328. Epub 2014 Jul 4. PMID: 25064373; PMCID: PMC4170722.
- Yuan S, Chen J, Ruan X, Sun Y, Zhang K, Wang X, Li X, Gill D, Burgess S, Giovannucci E, Larsson SC. Smoking, alcohol consumption, and 24 gastrointestinal diseases: Mendelian randomization analysis. Elife. 2023 Feb 2;12:e84051. doi: 10.7554/eLife.84051. PMID: 36727839; PMCID: PMC10017103.
- Xiong J, Yang L, Deng YQ, Yan SY, Gu JM, Li BH, Zi H, Ming DJ, Zeng XT, Wang YB. The causal association between smoking, alcohol consumption and risk of bladder cancer: A univariable and multivariable Mendelian randomization study. Int J Cancer. 2022 Dec 15;151(12):2136-2143. doi: 10.1002/ijc.34228. Epub 2022 Aug 11. PMID: 35904850.
- Shin SY, Fauman EB, Petersen AK, Krumsiek J, Santos R, Huang J, Arnold M, Erte I, Forgetta V, Yang TP, Walter K, Menni C, Chen L, Vasquez L, Valdes AM, Hyde CL, Wang V, Ziemek D, Roberts P, Xi L, Grundberg E; Multiple Tissue Human Expression Resource (MuTHER) Consortium; Waldenberger M, Richards JB, Mohney RP, Milburn MV, John SL, Trimmer J, Theis FJ, Overington JP, Suhre K, Brosnan MJ, Gieger C, Kastenmüller G, Spector TD, Soranzo N. An atlas of genetic influences on human blood metabolites. Nat Genet. 2014 Jun;46(6):543-550. doi: 10.1038/ng.2982. Epub 2014 May 11. PMID: 24816252; PMCID: PMC4064254.
- Sanna S, van Zuydam NR, Mahajan A, Kurilshikov A, Vich Vila A, Võsa U, Mujagic Z, Masclee AAM, Jonkers DMAE, Oosting M, Joosten LAB, Netea MG, Franke L, Zhernakova A, Fu J, Wijmenga C, McCarthy MI. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat Genet. 2019 Apr;51(4):600-605. doi: 10.1038/s41588-019-0350-x. Epub 2019 Feb 18. PMID: 30778224; PMCID: PMC6441384.
- Dudbridge F. Polygenic Mendelian Randomization. Cold Spring Harb Perspect Med. 2021 Feb 1;11(2):a039586. doi: 10.1101/cshperspect.a039586. PMID: 32229610; PMCID: PMC7849343.
- Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015 Apr;44(2):512-25. doi: 10.1093/ije/dyv080. Epub 2015 Jun 6. PMID: 26050253; PMCID: PMC4469799.
- Cohen JF, Chalumeau M, Cohen R, Korevaar DA, Khoshnood B, Bossuyt PM. Cochran's Q test was useful to assess heterogeneity in likelihood ratios in studies of diagnostic accuracy. J Clin Epidemiol. 2015 Mar;68(3):299-306. doi: 10.1016/j.jclinepi.2014.09.005. Epub 2014 Oct 23. PMID: 25441698.
- Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018 May;50(5):693-698. doi: 10.1038/s41588-018-0099-7. Epub 2018 Apr 23. Erratum in: Nat Genet. 2018 Aug;50(8):1196. PMID: 29686387; PMCID: PMC6083837.
- Nolte IM. Metasubtract: an R-package to analytically produce leave-one-out meta-analysis GWAS summary statistics. Bioinformatics. 2020 Aug 15;36(16):4521-4522. doi: 10.1093/bioinformatics/btaa570. PMID: 32696040; PMCID: PMC7750933.
- Lunjani N, Walsh LJ, Venter C, Power M, MacSharry J, Murphy DM, O'Mahony L. Environmental influences on childhood asthma-The effect of diet and microbiome on asthma. Pediatr Allergy Immunol. 2022 Dec;33(12):e13892. doi: 10.1111/pai.13892. PMID: 36564884; PMCID: PMC10107834.
- Kozik AJ, Holguin F, Segal LN, Chatila TA, Dixon AE, Gern JE, Lozupone C, Lukacs N, Lumeng C, Molyneaux PL, Reisdorph N, Vujkovic-Cvijin I, Togias A, Huang YJ. Microbiome, Metabolism, and Immunoregulation of Asthma: An American Thoracic Society and National Institute of Allergy and Infectious Diseases Workshop Report. Am J Respir Cell Mol Biol. 2022 Aug;67(2):155-163. doi: 10.1165/rcmb.2022-0216ST. PMID: 35914321; PMCID: PMC9348558.
- Farraia M, Cavaleiro Rufo J, Paciência I, Castro Mendes F, Delgado L, Laerte Boechat J, Moreira A. Metabolic interactions in asthma. Eur Ann Allergy Clin Immunol. 2019 Sep 16;51(5):196-205. doi: 10.23822/EurAnnACI.1764-1489.101. Epub 2019 Jul 9. PMID: 31287260.
- Kelly RS, Dahlin A, McGeachie MJ, Qiu W, Sordillo J, Wan ES, Wu AC, Lasky-Su J. Asthma Metabolomics and the Potential for Integrative Omics in Research and the Clinic. Chest. 2017 Feb;151(2):262-277. doi: 10.1016/j.chest.2016.10.008. Epub 2016 Oct 21. PMID: 27776981; PMCID: PMC5310123.
- Xu S, Panettieri RA Jr, Jude J. Metabolomics in asthma: A platform for discovery. Mol Aspects Med. 2022 Jun;85:100990. doi: 10.1016/j.mam.2021.100990. Epub 2021 Jul 17. PMID: 34281719; PMCID: PMC9088882.
- Jiang T, Dai L, Li P, Zhao J, Wang X, An L, Liu M, Wu S, Wang Y, Peng Y, Sun D, Zheng C, Wang T, Wen X, Cheng Z. Lipid metabolism and identification of biomarkers in asthma by lipidomic analysis. Biochim Biophys Acta Mol Cell Biol Lipids. 2021 Feb;1866(2):158853. doi: 10.1016/j.bbalip.2020.158853. Epub 2020 Nov 4. PMID: 33160078.
- Rodriguez-Perez N, Schiavi E, Frei R, Ferstl R, Wawrzyniak P, Smolinska S, Sokolowska M, Sievi NA, Kohler M, Schmid-Grendelmeier P, Michalovich D, Simpson KD, Hessel EM, Jutel M, Martin-Fontecha M, Palomares O, Akdis CA, O'Mahony L. Altered fatty acid metabolism and reduced stearoyl-coenzyme a desaturase activity in asthma. Allergy. 2017 Nov;72(11):1744-1752. doi: 10.1111/all.13180. Epub 2017 May 10. PMID: 28397284.
- Xing SC, Mi JD, Chen JY, Hu JX, Liao XD. Metabolic activity of Bacillus coagulans R11 and the health benefits of and potential pathogen inhibition by this species in the intestines of laying hens under lead exposure. Sci Total Environ. 2020 Mar 20;709:134507. doi: 10.1016/j.scitotenv.2019.134507. Epub 2019 Dec 16. PMID: 31881475.
- Morris SM Jr. Regulation of enzymes of the urea cycle and arginine metabolism. Annu Rev Nutr. 2002;22:87-105. doi: 10.1146/annurev.nutr.22.110801.140547. Epub 2002 Jan 4. PMID: 12055339.
- Nagagarajan S, Xue F, MacKerell AD Jr. Impact of substrate protonation and tautomerization states on interactions with the active site of arginase I. J Chem Inf Model. 2013 Feb 25;53(2):452-60. doi: 10.1021/ci300506y. Epub 2013 Jan 31. PMID: 23327293; PMCID: PMC3590304.
- Lara A, Khatri SB, Wang Z, Comhair SA, Xu W, Dweik RA, Bodine M, Levison BS, Hammel J, Bleecker E, Busse W, Calhoun WJ, Castro M, Chung KF, Curran-Everett D, Gaston B, Israel E, Jarjour N, Moore W, Peters SP, Teague WG, Wenzel S, Hazen SL, Erzurum SC; National Heart, Lung, and Blood Institute's Severe Asthma Research Program. Alterations of the arginine metabolome in asthma. Am J Respir Crit Care Med. 2008 Oct 1;178(7):673-81. doi: 10.1164/rccm.200710-1542OC. Epub 2008 Jul 17. Erratum in: Am J Respir Crit Care Med. 2012 Nov 1;186(9):932. PMID: 18635886; PMCID: PMC2556449.
- Coman D, Yaplito-Lee J, Boneh A. New indications and controversies in arginine therapy. Clin Nutr. 2008 Aug;27(4):489-96. doi: 10.1016/j.clnu.2008.05.007. Epub 2008 Jul 21. PMID: 18640748.
- Luo J, Chen H, Zhang Q, Huang X, Qin X, Li J, Chen S, Xiao Y, Sun L, Sun B. Metabolism Characteristics of Mycoplasma pneumoniae Infection in Asthmatic Children. Allergy Asthma Immunol Res. 2022 Nov;14(6):713-729. doi: 10.4168/aair.2022.14.6.713. PMID: 36426399; PMCID: PMC9709688.
- Bazzano M, Laghi L, Zhu C, Magi GE, Serri E, Spaterna A, Tesei B, Laus F. Metabolomics of tracheal wash samples and exhaled breath condensates in healthy horses and horses affected by equine asthma. J Breath Res. 2018 Sep 27;12(4):046015. doi: 10.1088/1752-7163/aade13. PMID: 30168442.
- Althoff MD, Jimenez G, Peterson R, Jin Y, Grasemann H, Sharma S, Federman AD, Wisnivesky JP, Holguin F. Differences in L-arginine metabolism and asthma morbidity among asthma patients with and without obstructive sleep apnea. Respir Res. 2022 Sep 5;23(1):230. doi: 10.1186/s12931-022-02157-9. PMID: 36064404; PMCID: PMC9442950.
- Althoff MD, Peterson R, McGrath M, Jin Y, Grasemann H, Sharma S, Federman A, Wisnivesky JP, Holguin F. Phenotypic characteristics of asthma and morbidity are associated with distinct longitudinal changes in L-arginine metabolism. BMJ Open Respir Res. 2023 Jun;10(1):e001683. doi: 10.1136/bmjresp-2023-001683. PMID: 37270184; PMCID: PMC10254613.
- Scott JA, Grasemann H. Arginine metabolism in asthma. Immunol Allergy Clin North Am. 2014 Nov;34(4):767-75. doi: 10.1016/j.iac.2014.07.007. Epub 2014 Aug 19. PMID: 25282289.
- Ludviksdottir D, Diamant Z, Alving K, Bjermer L, Malinovschi A. Clinical aspects of using exhaled NO in asthma diagnosis and management. Clin Respir J. 2012 Oct;6(4):193-207. doi: 10.1111/crj.12001. PMID: 22898078.
- Darwish AG, Das PR, Ismail A, Gajjar P, Balasubramani SP, Sheikh MB, Tsolova V, Sherif SM, El-Sharkawy I. Untargeted Metabolomics and Antioxidant Capacities of Muscadine Grape Genotypes during Berry Development. Antioxidants (Basel). 2021 Jun 4;10(6):914. doi: 10.3390/antiox10060914. PMID: 34200012; PMCID: PMC8230005.
- Thomson NC. The Role of Smoking in Asthma and Chronic Obstructive Pulmonary Disease Overlap. Immunol Allergy Clin North Am. 2022 Aug;42(3):615-630. doi: 10.1016/j.iac.2022.03.004. Epub 2022 Jun 30. PMID: 35965049.
- Yang W, Yang Y, He L, Zhang M, Sun S, Wang F, Han B. Dietary factors and risk for asthma: A Mendelian randomization analysis. Front Immunol. 2023 Feb 22;14:1126457. doi: 10.3389/fimmu.2023.1126457. PMID: 36911739; PMCID: PMC9992976.
- Mikkelsen H, Landt EM, Benn M, Nordestgaard BG, Dahl M. Causal risk factors for asthma in Mendelian randomization studies: A systematic review and meta-analysis. Clin Transl Allergy. 2022 Nov;12(11):e12207. doi: 10.1002/clt2.12207. PMID: 36434743; PMCID: PMC9640961.
- Sachdeva K, Do DC, Zhang Y, Hu X, Chen J, Gao P. Environmental Exposures and Asthma Development: Autophagy, Mitophagy, and Cellular Senescence. Front Immunol. 2019 Nov 29;10:2787. doi: 10.3389/fimmu.2019.02787. PMID: 31849968; PMCID: PMC6896909.
- Lu C, Zhang Y, Li B, Zhao Z, Huang C, Zhang X, Qian H, Wang J, Liu W, Sun Y, Norbäck D, Deng Q. Interaction effect of prenatal and postnatal exposure to ambient air pollution and temperature on childhood asthma. Environ Int. 2022 Sep;167:107456. doi: 10.1016/j.envint.2022.107456. Epub 2022 Aug 6. PMID: 35952466.